Introduction
The inferior colliculus (IC; plural: colliculi) is a paired structure in the midbrain, which serves as an important relay point for auditory information as it travels from the inner ear to the auditory cortex. It is responsible for integrated sound localization, as it is the first site at which input from cells carrying horizontal and vertical sound location data from each ear converge. Also, it plays an important role in generating the startle response, orienting the body toward relevant stimuli, and discriminating pitch and rhythm. It receives signals from many non-auditory sources, but primarily from the other brainstem nuclei involved in the auditory system. Efferent projections from the IC often travel to the many nuclei which send it afferents, but especially to the medial geniculate nucleus of the thalamus.[1][2][3]
Structure and Function
Register For Free And Read The Full Article
 Search engine and full access to all medical articles
Search engine and full access to all medical articles- 10 free questions in your specialty
- Free CME/CE Activities
- Free daily question in your email
- Save favorite articles to your dashboard
- Emails offering discounts
Learn more about a Subscription to StatPearls Point-of-Care
Structure and Function
The two ICs lie rostral to the trochlear nerve and just caudal to the superior colliculi. Their location is on the dorsal surface of the rostral midbrain, ventral to the lateral lemniscus. Each set of colliculi resembles two symmetrical bumps. The two sets of colliculi (superior and inferior) comprise the quadrigeminal cistern, part of the tectum or “roof” of the midbrain. [2][4]
Glutamatergic cells in layers V and VI of the auditory cortex project principally to the ipsilateral IC, with the majority of cortical input coming from layer V. This cortical input, allows for top-down modulation of IC response to frequency, intensity, duration, and location of auditory stimuli, and may play a role in discrimination of multiple different sound sources.[5] The IC contains mostly glutamatergic neurons, which receive and integrate auditory input from the cortex, medial geniculate nucleus, cerebellum, lateral lemniscus, and the contralateral IC. A minority of neurons are GABAergic and inhibit neurons in the medial geniculate body and contralateral IC. There is a hypothesis that they play a role in refining the auditory information conducted to higher structures. The major output from the IC runs through the brachium of the inferior colliculus to excite the medial geniculate nucleus of the thalamus. Another major source of output runs to the contralateral IC and lateral lemniscus.[2][6]
There are three regions of the IC: the central nucleus, the dorsal cortex, and the external cortex. The dorsal and external cortices are the major sites of non-auditory input to the IC, discussed below.[7] The central nucleus is so named because the other two regions encircle it; it is the largest nucleus and the major site of auditory inputs to the IC. The central nucleus sends tonotopically organized efferent fibers, ultimately to the primary auditory cortex in the temporal lobe.[8] Pitch location in the cochlea is also mappable within the central nucleus.[9] Discrimination between two different concurrent pitches is achieved in the IC and aided by the timing of their arrival at the IC.[10] Differences in the gray matter density and activity level of the IC modulate the perception and dislike of harmonic dissonance, likely related to pitch discrimination.[11][12] Interestingly, the IC pitch representation differs between those who speak tonal languages, such as Chinese, and those who speak non-tonal languages.[13] Cells in the IC are selectively active depending on the horizontal and vertical angle from which sound arrives at the ears of the listener and the rhythm of those sounds.[14][15][16]
Not only does the IC allow for sound localization by integrating the input from both ears, but it also receives many non-auditory inputs. Some cells in IC respond to contralateral visual field stimuli, sending and receiving information from the superior colliculi. While their purpose is unknown, the theory is that they help map physical space with both modalities of information. The auditory and visual inputs regarding where a sound originated are also joined by somatosensory inputs from the spinal cord and cranial nerves, which researchers hypothesize aid one’s ability to orient the body toward the source of the sound. When an unexpected or loud sound occurs, the IC helps to mediate the ensuing startle response. Additionally, the IC is more active when perceiving communication by a member of the same species, indicating a special interest in salient environmental noises.[3][17] When paying greater attention to a task (for example, in anticipation of a reward or during opposite-sex social encounters), the IC is more active. In contrast, the IC also plays a role in filtering out noises that are not important, such as self-generated chewing and breathing.[3][18] IC activity changes with body movement and posture, additional information that serves to discriminate between internal sounds and sounds in the environment.[7]
Embryology
The precursor to the brain and spinal cord, the neural tube, begins closing to form its cylindrical shape around embryonic day 21. The tube then begins to bend and expand to further specialized structures, in response to chemical signals from nearby tissues that influence their developmental fate. Those signals cause the neural tube to develop into three primary vesicles: the prosencephalon, mesencephalon, and rhombencephalon. By day 24, the neural tube further develops into secondary vesicles. The prosencephalon divides into telencephalon and diencephalon, the rhombencephalon develops into metencephalon and myelencephalon, and the mesencephalon does not further divide. The mesencephalon, also known as the midbrain, is the embryonic structure from which the tectum forms.[19] The dorsal portion of the mesencephalon is the portion from which derive the inferior and superior colliculi.[20]
Multiple molecular signals are essential in the process of IC formation, including fibroblast growth factor (FGF) 8 and FGF17. The absence of either of the growth factors mentioned earlier results in abnormal formation of the midbrain and cerebellum in mice.[20] There is also a prominent role of Pax2 and Pax5 proteins in the development of the midbrain in mice. Knockout of Pax2 in Zebrafish disrupts the organization of the midbrain, while knockout of Pax5 protein results in underdeveloped or absent IC.[21] While many more inductive signals are vital for the proper development of this structure, one final notable example is Neurod1, which is a protein essential for the tonotopic structure of the auditory system throughout the pathway— in the ear, cochlear nuclei, and IC.[22] Tonotopy of the IC continues to develop after birth, and upon the comparison between postnatal days 9 and 20 in mice, the frequency responsiveness and characteristic frequencies of different IC cell populations had changed and developed.[23]
Blood Supply and Lymphatics
The two vertebral arteries within the neck merge at the base of the skull to form one midline basilar artery. The blood supply of the medial IC mainly flows from the paramedian branches of the basilar artery. The lateral IC primarily receives vascular supply via the rostral trunk of the superior cerebellar artery, another branch arising from the basilar artery.[2][4] Additional blood supply to the IC comes from the collicular and posteromedial choroidal arteries, both proximal branches of the posterior cerebral artery (which also arises from the basilar artery).[24] Blockage to any of the more distal or proximal branches may impact the functioning of the inferior colliculus.
Previously, anatomists believed that the brain lacked lymphatic drainage. A growing body of evidence reveals the presence of a glymphatic system, which then may drain through the cerebrospinal fluid to the periphery.[25] Found to lie along the surface of much of the Zebrafish brain, brain lymphatic endothelial cells are closely associated with blood vessels.[26]
Nerves
The IC receives information from the lateral lemniscus, which contains neurons from the cochlear nuclei and superior olive relaying auditory information from the cochlear nerve.[4] The cochlear nerve is a portion of the vestibulocochlear nerve or CN VIII. It is responsible for carrying auditory signals from the cochlea, while the vestibular portion carries information from the semicircular canals about the position of the head.[27] In its auditory functions, such as pitch localization and recognition, the IC primarily utilizes neural input from the cochlear nerve. It also integrates efferent information mostly from the ipsilateral auditory cortex, referred to as cortico-collicular projections.[5]
However, the IC also receives input from and sends output to the retina and superior colliculi, which receive visual information through the optic nerve, CN II. As mentioned above, these connections may aid in the creation of a mental map of physical space. Additionally, multiple spinal somatosensory nerves, mostly from the dorsal column of the spinal cord and peripheral nerves, also input to the IC to allow coordination of movement in response to different behaviorally relevant auditory stimuli, including a startle response away from unexpected noises. The IC is sensitive to inputs from the trigeminal nerve, CN V, and its associated ganglion.[3]
Muscles
While many skeletal muscles will be involved in the ability to orient oneself to relevant stimuli and produce a startle response, the muscles that may be involved with IC functioning have not been fully elucidated.[3] The auditory pathway contains two small muscles in the middle ear: the tensor tympani and the stapedius. Both muscles play a role in stiffening the tympanic membrane when they contract, decreasing the amount of movement transmitted through the middle ear ossicles and thus lowering the perceived amplitude or volume of sounds.[28][29] In this way, they aid in preventing auditory damage due to loud noises, which have been shown to cause hearing loss and change the tonotopy, frequency thresholds, excitability, and firing rates of IC neurons specifically.[30]
Surgical Considerations
The IC lies near many important areas controlling vital functions, and caution is necessary during any surgical manipulations of the area. Most of the midbrain tracts and nuclear groups are located at the level of the IC, and thus lesions in this area can have widespread effects. One of the most frequent complications of tumors in the midbrain is hydrocephalus due to blockage of the cerebral aqueduct. Nothnagel syndrome, characterized by ipsilateral or bilateral oculomotor nerve paralysis and ipsilateral cerebellar ataxia, is another pathology associated with tumors of the midbrain.[2]
Surgery to the tectum may be considered as a treatment for tectal glioma, especially for those with neurofibromatosis-1, because these patients’ tumors tend to grow more often and at a faster rate.[31] One common complication of tumor resection and other surgical manipulations in the midbrain is Parinaud syndrome, characterized by issues with vertical gaze due to damage of the rostral interstitial nucleus of the medial longitudinal fasciculus.[2][32][33]
Another surgery in this area is the placement of an IC implant, which works similarly to a cochlear implant but is utilized in those with cochlear nerve damage. These implants may be used especially for those with neurofibromatosis-2 whose cochlear nerves suffer damage upon removal of schwannomas of CN VIII. A trans-labyrinthine or retrosigmoid approach with possible endoscopic aid may be the best approach to the area surgically, and brainstem auditory evoked potentials (discussed below) can be monitored during surgery to avoid damage to auditory pathways.[4][32]
Clinical Significance
As the level of the IC contains many important structures, IC lesions can result in several clinical syndromes. Lesions of the IC include tumors, hemorrhages, and other derangements. Damage in one IC has resulted in many auditory issues, such as tinnitus, hyperacusis, audiogenic seizures, auditory agnosia, deficits in contralateral hemispace sound localization, and difficulty recognizing speech among other concurrent noises on the ipsilateral side.[30][34][35][36][37] Damage to the IC can also result in signs which stem from nearby structures, such as contralateral hemisensory loss (spinothalamic tract disturbance) and vertical diplopia (trochlear nerve palsies).[36]
In animal models, deep brain stimulation of the external cortex of the IC results in reduced tinnitus, and the central nucleus has been implicated in the generation of tinnitus.[34][38] In human tinnitus patients, differences in morphology and gray matter volume in the IC, as well as changes in IC activation, have been observed.[39]
The IC may have some involvement in audiogenic seizures, as studies of genetically epilepsy-prone rats have revealed IC excitatory and inhibitory balance derangements, which may play a role in seizure initiation.[40][41] Damage to the central nucleus IC in mice has resulted in temporary deficits in the startle response, while dorsal and external nuclei lesions have resulted in increased startle response.[42] Further investigation in each of these areas will be required to determine their applicability to humans.
Auditory evoked potentials are useful for diagnosing pathologies in the auditory pathway as well as monitoring the auditory system during surgery. The waveform of the auditory evoked potentials corresponds to different portions of the pathway, with wave V (and the negative potential that follows) attributed to the activity of the lateral lemniscus and IC.[43][44] Lesions of the IC result in the reduction of those specific waves.[45]
Media
(Click Image to Enlarge)
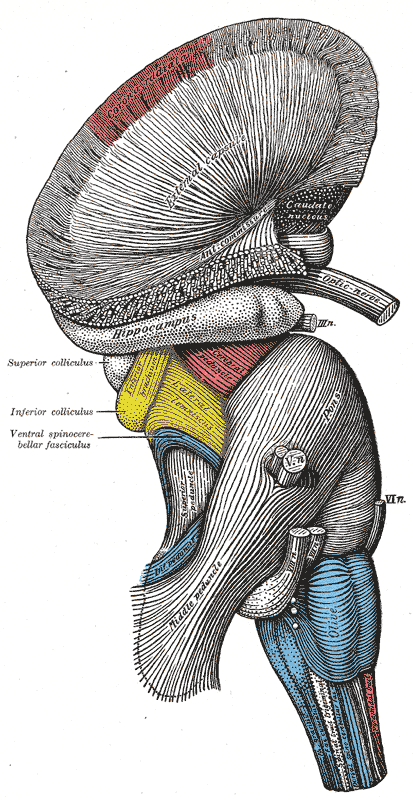
Hindbrain, Superficial Dissection. This lateral-view illustration shows the relationships between the hindbrain (rhombencephalon) structures. The structures included are the corona radiata, external capsule, anterior commissure, hippocampus, cerebral peduncle, superior and inferior colliculus, brachium of the inferior colliculus, lateral lemniscus, ventral spinocerebellar fasciculus, pons, superior, middle, and inferior peduncles, optic, oculomotor, trigeminal, abducens, facial, and vestibulocochlear nerves, olive, and medulla oblongata.
Henry Vandyke Carter, Public Domain, via Wikimedia Commons
(Click Image to Enlarge)
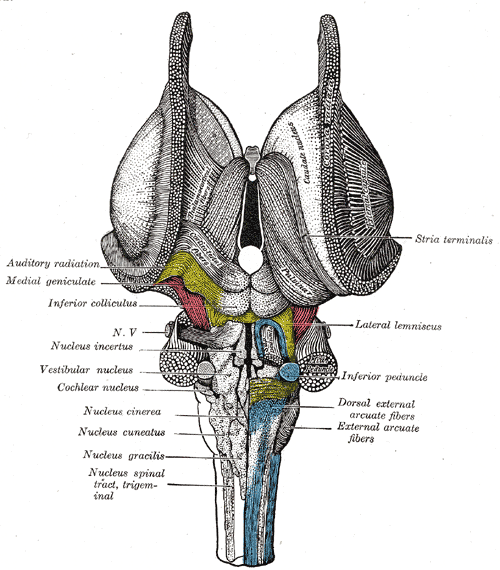
The Hindbrain or Rhombencephalon, Dorsal View. This is a dissection of the brain stem with a dorsal view of the stria terminalis, auditory radiation, medial geniculate, inferior colliculus, inferior peduncle nucleus cinerea, cuneatus, gracilis, nucleus spinal tract, and trigeminal.
Henry Vandyke Carter, Public Domain, via Wikimedia Commons
References
Mei HX, Cheng L, Chen QC. Neural interactions in unilateral colliculus and between bilateral colliculi modulate auditory signal processing. Frontiers in neural circuits. 2013:7():68. doi: 10.3389/fncir.2013.00068. Epub 2013 Apr 19 [PubMed PMID: 23626523]
Level 3 (low-level) evidenceRuchalski K, Hathout GM. A medley of midbrain maladies: a brief review of midbrain anatomy and syndromology for radiologists. Radiology research and practice. 2012:2012():258524. doi: 10.1155/2012/258524. Epub 2012 May 22 [PubMed PMID: 22693668]
Gruters KG, Groh JM. Sounds and beyond: multisensory and other non-auditory signals in the inferior colliculus. Frontiers in neural circuits. 2012:6():96. doi: 10.3389/fncir.2012.00096. Epub 2012 Dec 11 [PubMed PMID: 23248584]
Komune N, Yagmurlu K, Matsuo S, Miki K, Abe H, Rhoton AL Jr. Auditory brainstem implantation: anatomy and approaches. Neurosurgery. 2015 Jun:11 Suppl 2():306-20; discussion 320-1. doi: 10.1227/NEU.0000000000000736. Epub [PubMed PMID: 25793729]
Bajo VM, King AJ. Cortical modulation of auditory processing in the midbrain. Frontiers in neural circuits. 2012:6():114. doi: 10.3389/fncir.2012.00114. Epub 2013 Jan 3 [PubMed PMID: 23316140]
Ito T, Oliver DL. The basic circuit of the IC: tectothalamic neurons with different patterns of synaptic organization send different messages to the thalamus. Frontiers in neural circuits. 2012:6():48. doi: 10.3389/fncir.2012.00048. Epub 2012 Jul 26 [PubMed PMID: 22855671]
Yang Y, Lee J, Kim G. Integration of locomotion and auditory signals in the mouse inferior colliculus. eLife. 2020 Jan 28:9():. doi: 10.7554/eLife.52228. Epub 2020 Jan 28 [PubMed PMID: 31987070]
Straka MM, Hughes R, Lee P, Lim HH. Descending and tonotopic projection patterns from the auditory cortex to the inferior colliculus. Neuroscience. 2015 Aug 6:300():325-37. doi: 10.1016/j.neuroscience.2015.05.032. Epub 2015 May 19 [PubMed PMID: 26002315]
Merzenich MM, Reid MD. Representation of the cochlea within the inferior colliculus of the cat. Brain research. 1974 Sep 13:77(3):397-415 [PubMed PMID: 4854119]
Level 3 (low-level) evidenceChen C, Read HL, Escabí MA. A temporal integration mechanism enhances frequency selectivity of broadband inputs to inferior colliculus. PLoS biology. 2019 Jun:17(6):e2005861. doi: 10.1371/journal.pbio.2005861. Epub 2019 Jun 24 [PubMed PMID: 31233489]
Fritz TH, Renders W, Müller K, Schmude P, Leman M, Turner R, Villringer A. Anatomical differences in the human inferior colliculus relate to the perceived valence of musical consonance and dissonance. The European journal of neuroscience. 2013 Oct:38(7):3099-105. doi: 10.1111/ejn.12305. Epub 2013 Jul 16 [PubMed PMID: 23859464]
Kim SG, Lepsien J, Fritz TH, Mildner T, Mueller K. Dissonance encoding in human inferior colliculus covaries with individual differences in dislike of dissonant music. Scientific reports. 2017 Jul 18:7(1):5726. doi: 10.1038/s41598-017-06105-2. Epub 2017 Jul 18 [PubMed PMID: 28720776]
Krishnan A, Gandour JT. The role of the auditory brainstem in processing linguistically-relevant pitch patterns. Brain and language. 2009 Sep:110(3):135-48. doi: 10.1016/j.bandl.2009.03.005. Epub 2009 Apr 14 [PubMed PMID: 19366639]
Level 3 (low-level) evidenceAitkin L, Martin R. Neurons in the inferior colliculus of cats sensitive to sound-source elevation. Hearing research. 1990 Dec:50(1-2):97-105 [PubMed PMID: 2076986]
Level 3 (low-level) evidenceAkimov AG, Egorova MA, Ehret G. Spectral summation and facilitation in on- and off-responses for optimized representation of communication calls in mouse inferior colliculus. The European journal of neuroscience. 2017 Feb:45(3):440-459. doi: 10.1111/ejn.13488. Epub 2017 Jan 4 [PubMed PMID: 27891665]
Day ML, Delgutte B. Neural population encoding and decoding of sound source location across sound level in the rabbit inferior colliculus. Journal of neurophysiology. 2016 Jan 1:115(1):193-207. doi: 10.1152/jn.00643.2015. Epub 2015 Oct 21 [PubMed PMID: 26490292]
Cabrera G, Cavelli M, Lopez C, Rodriguez-Servetti Z, Vanini G, Chase MH, Falconi A, Torterolo P. Wakefulness-promoting role of the inferior colliculus. Behavioural brain research. 2013 Nov 1:256():82-94. doi: 10.1016/j.bbr.2013.07.049. Epub 2013 Aug 7 [PubMed PMID: 23933516]
Level 3 (low-level) evidenceHurley LM, Sullivan MR. From behavioral context to receptors: serotonergic modulatory pathways in the IC. Frontiers in neural circuits. 2012:6():58. doi: 10.3389/fncir.2012.00058. Epub 2012 Sep 6 [PubMed PMID: 22973195]
Darnell D, Gilbert SF. Neuroembryology. Wiley interdisciplinary reviews. Developmental biology. 2017 Jan:6(1):. doi: 10.1002/wdev.215. Epub 2016 Dec 1 [PubMed PMID: 27906497]
Basson MA, Echevarria D, Ahn CP, Sudarov A, Joyner AL, Mason IJ, Martinez S, Martin GR. Specific regions within the embryonic midbrain and cerebellum require different levels of FGF signaling during development. Development (Cambridge, England). 2008 Mar:135(5):889-98. doi: 10.1242/dev.011569. Epub 2008 Jan 23 [PubMed PMID: 18216176]
Level 3 (low-level) evidenceUrbánek P, Fetka I, Meisler MH, Busslinger M. Cooperation of Pax2 and Pax5 in midbrain and cerebellum development. Proceedings of the National Academy of Sciences of the United States of America. 1997 May 27:94(11):5703-8 [PubMed PMID: 9159136]
Level 3 (low-level) evidenceMacova I, Pysanenko K, Chumak T, Dvorakova M, Bohuslavova R, Syka J, Fritzsch B, Pavlinkova G. Neurod1 Is Essential for the Primary Tonotopic Organization and Related Auditory Information Processing in the Midbrain. The Journal of neuroscience : the official journal of the Society for Neuroscience. 2019 Feb 6:39(6):984-1004. doi: 10.1523/JNEUROSCI.2557-18.2018. Epub 2018 Dec 12 [PubMed PMID: 30541910]
Romand R, Ehret G. Development of tonotopy in the inferior colliculus. I. Electrophysiological mapping in house mice. Brain research. Developmental brain research. 1990 Jul 1:54(2):221-34 [PubMed PMID: 2397588]
Level 3 (low-level) evidenceTatu L, Moulin T, Bogousslavsky J, Duvernoy H. Arterial territories of human brain: brainstem and cerebellum. Neurology. 1996 Nov:47(5):1125-35 [PubMed PMID: 8909417]
Louveau A, Smirnov I, Keyes TJ, Eccles JD, Rouhani SJ, Peske JD, Derecki NC, Castle D, Mandell JW, Lee KS, Harris TH, Kipnis J. Structural and functional features of central nervous system lymphatic vessels. Nature. 2015 Jul 16:523(7560):337-41. doi: 10.1038/nature14432. Epub 2015 Jun 1 [PubMed PMID: 26030524]
van Lessen M, Shibata-Germanos S, van Impel A, Hawkins TA, Rihel J, Schulte-Merker S. Intracellular uptake of macromolecules by brain lymphatic endothelial cells during zebrafish embryonic development. eLife. 2017 May 12:6():. pii: e25932. doi: 10.7554/eLife.25932. Epub 2017 May 12 [PubMed PMID: 28498105]
Moore JK, Perazzo LM, Braun A. Time course of axonal myelination in the human brainstem auditory pathway. Hearing research. 1995 Jul:87(1-2):21-31 [PubMed PMID: 8567438]
Keidar E, De Jong R, Kwartowitz G. Tensor Tympani Syndrome. StatPearls. 2023 Jan:(): [PubMed PMID: 30085597]
Dulak D, Naqvi IA. Neuroanatomy, Cranial Nerve 7 (Facial). StatPearls. 2023 Jan:(): [PubMed PMID: 30252375]
Niu Y, Kumaraguru A, Wang R, Sun W. Hyperexcitability of inferior colliculus neurons caused by acute noise exposure. Journal of neuroscience research. 2013 Feb:91(2):292-9. doi: 10.1002/jnr.23152. Epub 2012 Nov 14 [PubMed PMID: 23151900]
Level 3 (low-level) evidenceKershenovich A, Silman Z, de Rungs D, Koral K, Gargan L, Weprin B. Tectal Lesions in Children: A Long-Term Follow-Up Volumetric Tumor Growth Analysis in Surgical and Nonsurgical Cases. Pediatric neurosurgery. 2016:51(2):69-78. doi: 10.1159/000442795. Epub 2016 Jan 21 [PubMed PMID: 26789512]
Level 3 (low-level) evidenceLapras C, Bognar L, Turjman F, Villanyi E, Mottolese C, Fischer C, Jouvet A, Guyotat J. Tectal plate gliomas. Part I: Microsurgery of the tectal plate gliomas. Acta neurochirurgica. 1994:126(2-4):76-83 [PubMed PMID: 8042559]
Feroze KB, Patel BC. Parinaud Syndrome. StatPearls. 2023 Jan:(): [PubMed PMID: 28722922]
Kwee IL, Matsuzawa H, Nakada K, Fujii Y, Nakada T. Inferior colliculus syndrome: Clinical magnetic resonance microscopy anatomic analysis on a 7 T system. SAGE open medical case reports. 2017:5():2050313X17745209. doi: 10.1177/2050313X17745209. Epub 2017 Dec 5 [PubMed PMID: 29238582]
Level 3 (low-level) evidenceLitovsky RY, Fligor BJ, Tramo MJ. Functional role of the human inferior colliculus in binaural hearing. Hearing research. 2002 Mar:165(1-2):177-88 [PubMed PMID: 12031527]
Level 3 (low-level) evidenceStimmer H, Borrmann A, Löer C, Arnold W, Rummeny EJ. Monaural tinnitus from a contralateral inferior colliculus hemorrhage. Audiology & neuro-otology. 2009:14(1):35-8. doi: 10.1159/000152854. Epub 2008 Sep 4 [PubMed PMID: 18769025]
Level 3 (low-level) evidencePoliva O, Bestelmeyer PE, Hall M, Bultitude JH, Koller K, Rafal RD. Functional Mapping of the Human Auditory Cortex: fMRI Investigation of a Patient with Auditory Agnosia from Trauma to the Inferior Colliculus. Cognitive and behavioral neurology : official journal of the Society for Behavioral and Cognitive Neurology. 2015 Sep:28(3):160-80. doi: 10.1097/WNN.0000000000000072. Epub [PubMed PMID: 26413744]
Smit JV, Janssen ML, van Zwieten G, Jahanshahi A, Temel Y, Stokroos RJ. Deep brain stimulation of the inferior colliculus in the rodent suppresses tinnitus. Brain research. 2016 Nov 1:1650():118-124. doi: 10.1016/j.brainres.2016.08.046. Epub 2016 Aug 31 [PubMed PMID: 27592136]
Berger JI, Coomber B. Tinnitus-related changes in the inferior colliculus. Frontiers in neurology. 2015:6():61. doi: 10.3389/fneur.2015.00061. Epub 2015 Mar 30 [PubMed PMID: 25870582]
Ribak CE. An abnormal GABAergic system in the inferior colliculus provides a basis for audiogenic seizures in genetically epilepsy-prone rats. Epilepsy & behavior : E&B. 2017 Jun:71(Pt B):160-164. doi: 10.1016/j.yebeh.2015.02.024. Epub 2015 Mar 23 [PubMed PMID: 25812940]
Level 3 (low-level) evidenceFaingold CL, Naritoku DK, Copley CA, Randall ME, Riaz A, Anderson CA, Arnerić SP. Glutamate in the inferior colliculus plays a critical role in audiogenic seizure initiation. Epilepsy research. 1992 Nov:13(2):95-105 [PubMed PMID: 1361165]
Level 3 (low-level) evidenceParham K, Willott JF. Effects of inferior colliculus lesions on the acoustic startle response. Behavioral neuroscience. 1990 Dec:104(6):831-40 [PubMed PMID: 2285482]
Level 3 (low-level) evidencePlourde G. Auditory evoked potentials. Best practice & research. Clinical anaesthesiology. 2006 Mar:20(1):129-39 [PubMed PMID: 16634420]
Martin WH, Stecker MM. ASNM position statement: intraoperative monitoring of auditory evoked potentials. Journal of clinical monitoring and computing. 2008 Feb:22(1):75-85 [PubMed PMID: 18058246]
Level 1 (high-level) evidenceCaird DM, Klinke R. The effect of inferior colliculus lesions on auditory evoked potentials. Electroencephalography and clinical neurophysiology. 1987 May:68(3):237-40 [PubMed PMID: 2436886]
Level 3 (low-level) evidence