Introduction
The brain ranks as the most complex organ in the human body. The brain constantly receives numerous visual, auditory, olfactory, vestibular, proprioceptive, tactile, and gustatory sensory inputs. In addition to identifying and processing important information from these various sensory inputs, humans have a unique ability to suppress ruminative and socially unwanted behaviors. Research has attributed this function primarily to the prefrontal association cortex (PFC). Studies show that the PFC, mainly the dorsolateral prefrontal cortex, downregulates hippocampal activity to suppress unwanted thoughts.[1][2] The prefrontal association cortex is a cortical region in the anterior part of the cerebrum; it is involved in the top-down processing of sensory and motor information.[3]
The PFC is regarded as the center of higher cortical functions. Credit for significant knowledge of the PFC comes from the case of Phineas Gage, a railroad construction foreman whose personality changed entirely after a construction accident in the mid-19th century. Gage's PFC was destroyed when a rod pierced through his frontal lobe.
Once considered to be a thoughtful and decent man, he became a person who displayed socially inappropriate behavior due to the loss of the ability by the PFC to suppress unwanted behaviors. The knowledge of the prefrontal association cortex also carries implications for treating many psychiatric disorders, such as schizophrenia, major depressive disorders, and obsessive-compulsive disorders.[4]
Structure and Function
Register For Free And Read The Full Article
 Search engine and full access to all medical articles
Search engine and full access to all medical articles- 10 free questions in your specialty
- Free CME/CE Activities
- Free daily question in your email
- Save favorite articles to your dashboard
- Emails offering discounts
Learn more about a Subscription to StatPearls Point-of-Care
Structure and Function
The prefrontal association cortex is located in the anterior portion of the cerebral cortex, just anterior to the primary motor cortex and next to the premotor cortex. The prefrontal association cortex is bounded posteriorly by the Sylvian fissure. The PFC has significant networks of connections that run to and from sensory association cortices, the limbic system, the cortical regions of other lobes, and the dorsomedial nucleus of the thalamus. The prefrontal association cortex functions as an integration center for these multiple somatosensory inputs. The PFC is involved in memory formation, planning, execution, higher-order information processing, and suppression of unwanted behaviors. Interestingly, several studies have also shown the significance of the PFC in regulating endurance exercises, as evidenced by increased brain activity in this area with functional imaging.[5][6]
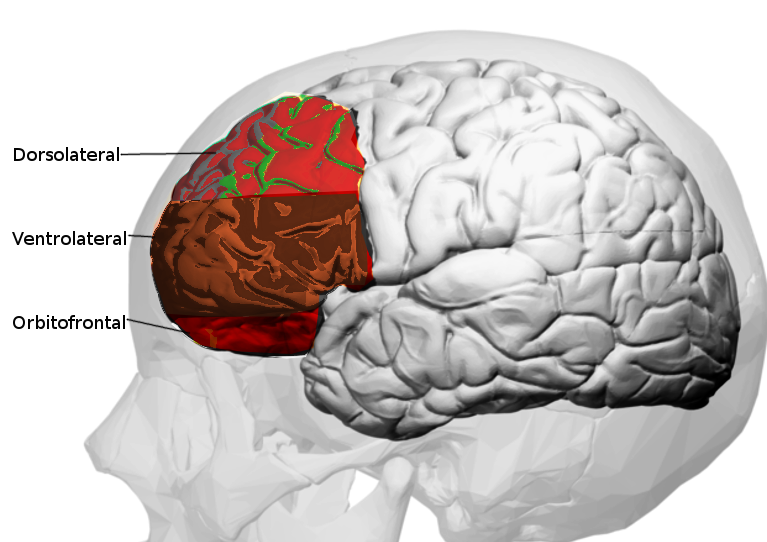
The PFC appears to subdivide into three different regions, namely, the dorsal, medial, and ventral prefrontal cortices. The dorsal PFC is the superior part; the medial PFC is found surrounding the principal sulcus, and the ventral PFC is found inferiorly around the orbit. The dorsolateral prefrontal cortex is involved in higher-order processing, whereas the ventral and medial prefrontal cortices play a part in regulating emotions. The medial and ventral prefrontal cortices communicate directly with the structures of the limbic system, such as the amygdala and cingulate cortex. Thus this area is involved in the regulation of emotion and memory.[2][7]
Anatomically, the medial PFC includes the anterior cingulate cortices (Brodmann area 24) and the subgenual cingulate cortex (Brodmann area 25). These areas are implicated in neuropathic pains, as evidenced by their hyperactivity during these events. In some instances, they were surgically removed as a treatment for the pain. The anterior cingulate cortex projects to adjacent premotor areas, such as Brodmann area 6d, and is involved in motor responses, such as eye and hand movements. The medial PFC also functions as the main visceromotor output for the PFC. Of note, the motor functions of the prefrontal association cortex are the result of projections from Brodmann area 25 to the subthalamic nucleus. Additionally, the PFC has significant projections to limbic areas, the ventral striatum, and the hypothalamus.[8][9]
Embryology
Human brain development starts during the third week of gestation and continues throughout early childhood and late adolescence. The ectoderm, one of the three germ layers of the embryo, contributes to the development of the nervous system. The notochord and somites, although not directly involved in the formation of the nervous system, play a critical role in the migration and patterning involved in the development of the nervous system. During the third week of gestation, the formation of the neural plate marks the beginning of the development of the nervous system. Through neurulation, neural plate formation begins a foundation for developing the neural tube. Along the neural plate midline, the neural groove forms.[8][10]
On either side of each groove, a structure called neural fold forms. This groove continues to deepen until week 4. At the same time, the neural folds start to fuse, which extends both rostrally and caudally. At this stage, the neural groove becomes a neural tube, leaving the two ends of the tube open (neuropores). The brain continues to develop from the neural tube. The primary brain vesicles (prosencephalon, mesencephalon, and rhombencephalon) form from the rostral neural tube. The primary vesicles continue to develop into different parts of the brain. Prosencephalon becomes telencephalon, eventually becoming adult structures such as the cerebral cortex, amygdala, pituitary, basal ganglia, and hippocampus. By the end of the eighth week of gestation, the foundation of the basic structures of the brain and CNS have formed.[8][10]
The PFC is among the last structures to fully develop. During the early embryonic stages of growth and development, the cells of the PFC characteristically demonstrate significant proliferation, followed by synapse elimination later in the stages of development. The extensive connectivity during early childhood undergoes pruning due to environmental interactions. Although the size of the brain approaches adult size by age 6, the cortex continues to develop through adulthood and adolescence.[10]
Blood Supply and Lymphatics
The blood supply to the prefrontal association cortex comes from the anterior circulation of the brain. The anterior circulation of the brain forms from the internal carotid artery and its branches, the anterior cerebral artery (ACA), and the middle cerebral artery (MCA). While the ACA supplies the middle and superior parts of the prefrontal association cortex, the MCA supplies the lateral and anterior portions.
The basal surface of the frontal lobe, the superior portion of the frontal gyrus, and a major portion of the precentral, central, and postcentral gyrus receive vascular supply from the cortical branches of the callosomarginal (a branch of the ACA) and distal peri-callosal artery. A superficial cerebral vein is the main vein drainage of the PFC, which then drains into the superior and inferior sagittal sinuses. Recent studies show that meningeal lymphatic vessels function as the lymphatic drainage of the brain.[11][12]
Nerves
There is no specific nerve supply to the prefrontal association cortex. Instead, the nerve supply to the prefrontal association cortex is explainable by numerous interconnections and projections to different parts of the brain. The PFC has reciprocal interconnections to various regions of the brain, such as the cortex, subcortical, and other brainstem regions, through bundles of axons. Association fibers (short and long fibers) form connections between different areas of the PFC and the other regions of the brain.
Several fasciculi, including the superior fronto-occipital fasciculus and inferior longitudinal fasciculus, form interconnections between the prefrontal association cortex and other brain areas. These fasciculi are involved in memory, languages, peripheral vision, object recognition, self-regulation, semantic processing, motivation, and emotion. These multi-synaptic connections between the prefrontal association cortex and anterior cingulate gyrus, amygdala, and mediodorsal thalamic nucleus mediate the inhibition of aggression and unwanted behavior.[13][14]
Physiologic Variants
Observations of physiologic variants of the prefrontal association cortex mainly occur in individuals exposed to stressors during the initial stages of growth and development. Studies have demonstrated a negative association between cumulative stress and the size of the PFC. Individuals with greater cumulative life stress tend to have smaller PFC, both in white and gray matter.
Introducing stressful events in the early stages of development impairs cognitive skills such as executive functions and spatial memory. Physiologic variations of the PFC also carry implications in neuropsychiatric disorders. Researchers also observed neuroanatomical differences in the PFC among individuals with genetic conditions such as 22q11.2 deletion syndrome.[15][16]
Surgical Considerations
Deep brain stimulation is an emerging novel therapeutic approach for treating neuropsychiatric disorders. Studies have demonstrated that deep brain stimulation of the internal capsule/striatum improves the function of the PFC. As a result, deep brain stimulation can be useful in treating neuropsychiatric disorders such as major depression and obsessive-compulsive disorders. The knowledge of the differences in the surface anatomy of the PFC in some genetic conditions is useful when considering surgical intervention. Since the PFC is bounded by different areas that can be tested and mapped, knowledge of this association cortex is important during surgeries to localize defects.[17][18]
Clinical Significance
The prefrontal association cortex correlates with various clinical cases and conditions such as major depressive disorders, substance abuse disorders, autism, Parkinson disease, prefrontal syndrome, schizophrenia, and obsessive-compulsive disorders. In individuals with major depressive disorders, the medial prefrontal cortex tends to be underactive or exhibits dysregulation. The subgenual cingulate cortex (Brodmann area 25) is thought to be overactive in depression. Thus, the PFC is the focal area for deep brain stimulation during treatment for depression.[3][7]
Injuries to the PFC result in disinhibition and disregard for social norms and values. A prefrontal syndrome is a condition that results from an injury or damage to the dorsal prefrontal association area. The dorsolateral part of the prefrontal cortex and the cingulate cortex are involved in memory, learning, cognitive function, and attention. Studies have depicted the presence of dysfunctions in motor activities with lesions of the PFC, such as impaired motor perseveration. Individuals with social anxiety disorder showed altered resting-state regional metabolism of the brain, such as in the medial dorsal prefrontal cortex, caudate, insula, and postcentral gyrus.[7][19]
Stress tends to reduce the firing rate of the PFC by weakening connectivity due to the opening of the potassium channel through the activation of cAMP pathways. Several studies on sleep physiology have demonstrated that the PFC is an area of origin for slow waves during the non-REM sleep cycle, as evidenced by the decreased regional cerebral blood flow to prefrontal areas during non-REM sleep.[3][7][19][20]
Media
(Click Image to Enlarge)
Prefrontal cortex Image courtesy O.Chaigasame
References
Schmitz TW, Correia MM, Ferreira CS, Prescot AP, Anderson MC. Hippocampal GABA enables inhibitory control over unwanted thoughts. Nature communications. 2017 Nov 3:8(1):1311. doi: 10.1038/s41467-017-00956-z. Epub 2017 Nov 3 [PubMed PMID: 29101315]
De Raedt R, Remue J, Loeys T, Hooley JM, Baeken C. The effect of transcranial direct current stimulation of the prefrontal cortex on implicit self-esteem is mediated by rumination after criticism. Behaviour research and therapy. 2017 Dec:99():138-146. doi: 10.1016/j.brat.2017.10.009. Epub 2017 Oct 21 [PubMed PMID: 29101842]
Datta D, Arnsten AFT. Loss of Prefrontal Cortical Higher Cognition with Uncontrollable Stress: Molecular Mechanisms, Changes with Age, and Relevance to Treatment. Brain sciences. 2019 May 17:9(5):. doi: 10.3390/brainsci9050113. Epub 2019 May 17 [PubMed PMID: 31108855]
Agarwal P, Sarris CE, Herschman Y, Agarwal N, Mammis A. Schizophrenia and neurosurgery: A dark past with hope of a brighter future. Journal of clinical neuroscience : official journal of the Neurosurgical Society of Australasia. 2016 Dec:34():53-58. doi: 10.1016/j.jocn.2016.08.009. Epub 2016 Sep 12 [PubMed PMID: 27634495]
Radel R, Tempest G, Denis G, Besson P, Zory R. Extending the limits of force endurance: Stimulation of the motor or the frontal cortex? Cortex; a journal devoted to the study of the nervous system and behavior. 2017 Dec:97():96-108. doi: 10.1016/j.cortex.2017.09.026. Epub 2017 Oct 9 [PubMed PMID: 29101820]
Kubera KM, Hirjak D, Wolf ND, Sambataro F, Thomann PA, Wolf RC. Intrinsic Network Connectivity Patterns Underlying Specific Dimensions of Impulsiveness in Healthy Young Adults. Brain topography. 2018 May:31(3):477-487. doi: 10.1007/s10548-017-0604-9. Epub 2017 Nov 3 [PubMed PMID: 29101492]
Callaghan BL, Dandash O, Simmons JG, Schwartz O, Byrne ML, Sheeber L, Allen NB, Whittle S. Amygdala Resting Connectivity Mediates Association Between Maternal Aggression and Adolescent Major Depression: A 7-Year Longitudinal Study. Journal of the American Academy of Child and Adolescent Psychiatry. 2017 Nov:56(11):983-991.e3. doi: 10.1016/j.jaac.2017.09.415. Epub 2017 Sep 19 [PubMed PMID: 29096781]
Selemon LD, Zecevic N. Schizophrenia: a tale of two critical periods for prefrontal cortical development. Translational psychiatry. 2015 Aug 18:5(8):e623. doi: 10.1038/tp.2015.115. Epub 2015 Aug 18 [PubMed PMID: 26285133]
Hathaway WR, Newton BW. Neuroanatomy, Prefrontal Cortex. StatPearls. 2023 Jan:(): [PubMed PMID: 29763094]
Stiles J, Jernigan TL. The basics of brain development. Neuropsychology review. 2010 Dec:20(4):327-48. doi: 10.1007/s11065-010-9148-4. Epub 2010 Nov 3 [PubMed PMID: 21042938]
Level 3 (low-level) evidenceKonan LM, Reddy V, Mesfin FB. Neuroanatomy, Cerebral Blood Supply. StatPearls. 2023 Jan:(): [PubMed PMID: 30335330]
Noé FM, Marchi N. Central nervous system lymphatic unit, immunity, and epilepsy: Is there a link? Epilepsia open. 2019 Mar:4(1):30-39. doi: 10.1002/epi4.12302. Epub 2019 Feb 14 [PubMed PMID: 30868113]
Schulz R, Runge CG, Bönstrup M, Cheng B, Gerloff C, Thomalla G, Hummel FC. Prefrontal-Premotor Pathways and Motor Output in Well-Recovered Stroke Patients. Frontiers in neurology. 2019:10():105. doi: 10.3389/fneur.2019.00105. Epub 2019 Feb 14 [PubMed PMID: 30837935]
Henschke JU, Oelschlegel AM, Angenstein F, Ohl FW, Goldschmidt J, Kanold PO, Budinger E. Early sensory experience influences the development of multisensory thalamocortical and intracortical connections of primary sensory cortices. Brain structure & function. 2018 Apr:223(3):1165-1190. doi: 10.1007/s00429-017-1549-1. Epub 2017 Nov 1 [PubMed PMID: 29094306]
Hanson JL, Chung MK, Avants BB, Rudolph KD, Shirtcliff EA, Gee JC, Davidson RJ, Pollak SD. Structural variations in prefrontal cortex mediate the relationship between early childhood stress and spatial working memory. The Journal of neuroscience : the official journal of the Society for Neuroscience. 2012 Jun 6:32(23):7917-25. doi: 10.1523/JNEUROSCI.0307-12.2012. Epub [PubMed PMID: 22674267]
Gudbrandsen M, Daly E, Murphy CM, Wichers RH, Stoencheva V, Perry E, Andrews D, Blackmore CE, Rogdaki M, Kushan L, Bearden CE, Murphy DGM, Craig MC, Ecker C. The Neuroanatomy of Autism Spectrum Disorder Symptomatology in 22q11.2 Deletion Syndrome. Cerebral cortex (New York, N.Y. : 1991). 2019 Jul 22:29(8):3655-3665. doi: 10.1093/cercor/bhy239. Epub [PubMed PMID: 30272146]
Widge AS, Zorowitz S, Basu I, Paulk AC, Cash SS, Eskandar EN, Deckersbach T, Miller EK, Dougherty DD. Deep brain stimulation of the internal capsule enhances human cognitive control and prefrontal cortex function. Nature communications. 2019 Apr 4:10(1):1536. doi: 10.1038/s41467-019-09557-4. Epub 2019 Apr 4 [PubMed PMID: 30948727]
Nejad AB, Rotgé JY, Valabregue R, Guérin-Langlois C, Hoertel N, Gorwood P, Dubertret C, Limosin F, Fossati P, Lemogne C. Medial prefrontal disengagement during self-focus in formerly depressed patients prone to rumination. Journal of affective disorders. 2019 Mar 15:247():36-44. doi: 10.1016/j.jad.2019.01.004. Epub 2019 Jan 4 [PubMed PMID: 30641339]
Doruyter A, Dupont P, Taljaard L, Stein DJ, Lochner C, Warwick JM. Resting regional brain metabolism in social anxiety disorder and the effect of moclobemide therapy. Metabolic brain disease. 2018 Apr:33(2):569-581. doi: 10.1007/s11011-017-0145-7. Epub 2017 Nov 3 [PubMed PMID: 29101601]
Tüshaus L, Omlin X, Tuura RO, Federspiel A, Luechinger R, Staempfli P, Koenig T, Achermann P. In human non-REM sleep, more slow-wave activity leads to less blood flow in the prefrontal cortex. Scientific reports. 2017 Nov 3:7(1):14993. doi: 10.1038/s41598-017-12890-7. Epub 2017 Nov 3 [PubMed PMID: 29101338]