Introduction
The dentate nucleus is the largest deep cerebellar cluster of neurons; it has a dentated – serrated – edge. Efferent fibers of the dentate nucleus are involved in the modulation of motor neurons and neurons involved in conscious thought and visuospatial function. Disruption of the dentate nucleus is typically associated with cerebellar ataxia.
Structure and Function
Register For Free And Read The Full Article
 Search engine and full access to all medical articles
Search engine and full access to all medical articles- 10 free questions in your specialty
- Free CME/CE Activities
- Free daily question in your email
- Save favorite articles to your dashboard
- Emails offering discounts
Learn more about a Subscription to StatPearls Point-of-Care
Structure and Function
The dentate nucleus is the most lateral deep cerebellar nucleus and is rich in iron. The cerebellar vermis and roof of the fourth ventricle are adjacent to the dentate nucleus. A sagittal section through the dentate nucleus shows its serrated appearance.[1]
The dentate nucleus regulates fine-control of voluntary movements, cognition, language, and sensory functions.
The dentate nucleus has two parts [2]:
- The dorsal (motor) domain: This has connections with cortical motor areas regulating motor functions
- The ventral (non-motor) domain: This has links with the cortical association areas controlling cognitive functions
Embryology
The cerebellum and, by extension, the dentate nucleus arises from the metencephalon, which derives from the rhombencephalon (hindbrain).[3] The cerebellar cells develop from the dorsal rhombomere 1.[4] The neurons that form the dentate nucleus and other deep cerebellar nuclei arise from the rostral rhombic lip, while GABAnergic cerebellar neurons such as Purkinje cells arise from the ventricular zone.[5][6] Dorsal rhombomere 1 is notable for the absence of Otx and Hox gene expression. Neuroblasts migrate from the rostral rhombic lip to the midline of the cerebellar primordium, then toward the nuclear transitory zone. From the nuclear transitory zone, the dentate nucleus neuroblasts move radially and express Pax6/Reelin, becoming Tbr2 positive and beginning formation of the dentate nucleus. Reelin is an extracellular glycoprotein important in the development of the brain, especially in the formation of the Purkinje cell layer.[7]
Fibroblast growth factor 8 (FGF8) is important in cerebellum development, as atopic injections of FGF8 can develop a secondary cerebellum in other brain regions. FGF8 appears to define the boundaries into which the cerebellum will develop. The subsequent removal of FGF8 establishes the onset of cerebellar development.[4]
Blood Supply and Lymphatics
The dentate nucleus mainly receives blood supply from the superior cerebellar artery (SCA) with some contribution from the posterior inferior cerebellar artery (PICA).[8] The SCA often supplies the dorsal, rostral, and lateral portions of the dentate nucleus while the PICA supplies the ventral and medial portions of the dentate nucleus. Neuroimaging suggests that the SCA supplies the motor dentate nucleus while the PICA supplies the nonmotor dentate nucleus.
Cerebrospinal fluid (CSF) derived from blood vessels supplying the dentate nucleus commonly comes from the PICA and the adjacent choroid plexus in the fourth ventricle.[9] The CSF enters circulation at the fourth ventricle and exits circulation within the subarachnoid space. Arachnoid villi transfer CSF from the subarachnoid space into veins and lymphatic vessels at the sheaths of the cranial and spinal nerves, the cribriform plate, and the adventitia of cerebral arteries.[10]
Nerves
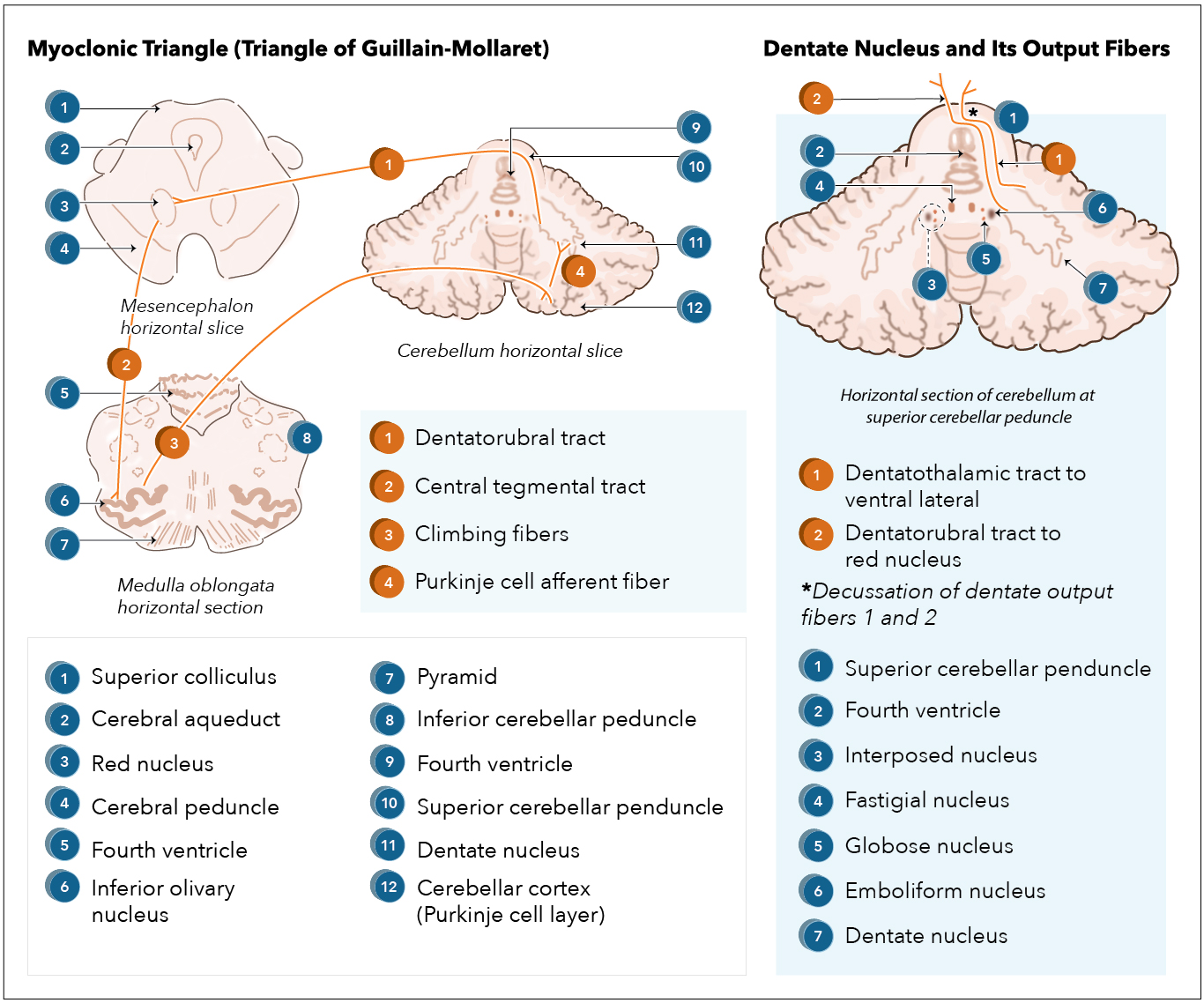
Inputs to the dentate nucleus. Afferent proprioceptive information synapses with dentate nucleus neurons through the spinocerebellar tract via the inferior cerebellar peduncle. Premotor cortex and supplementary cortex neurons also synapse the dentate nucleus.[1]
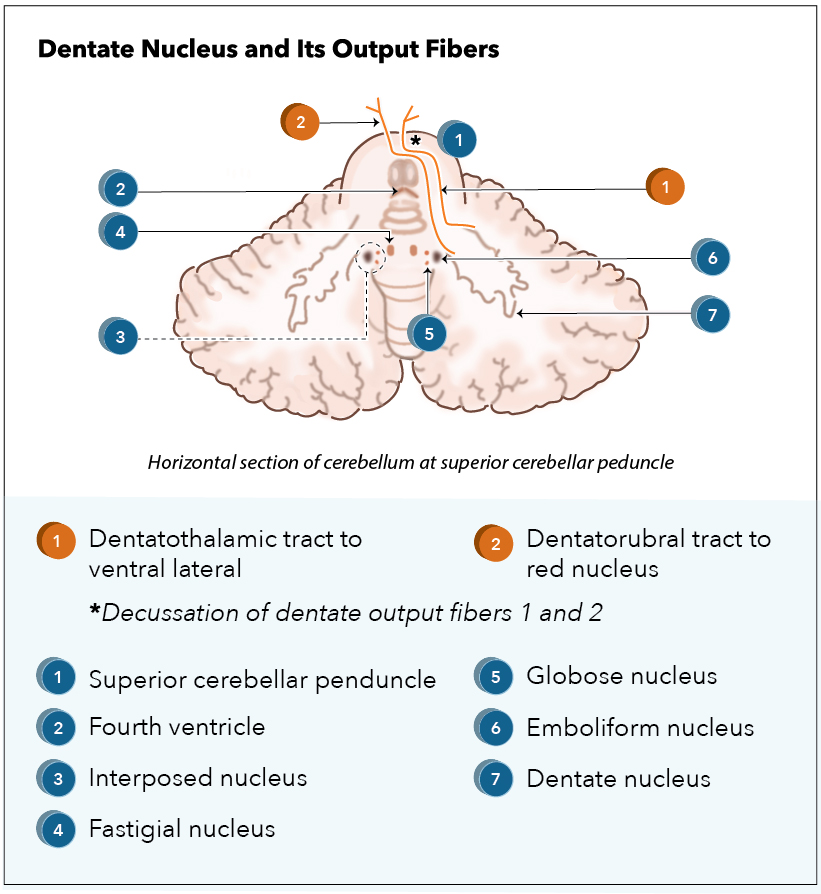
Outputs to the thalamus. Utilizing the dentatothalamic tract, the dentate nucleus sends output signals through the ipsilateral superior cerebral peduncle, then decussate to synapse the contralateral ventrolateral (VL) thalamic nucleus. VL neurons send fibers to the precentral gyrus (primary motor cortex; M1), premotor cortex, prefrontal gyri, posterior parietal areas, and the basal ganglia – specifically the striatum and the substantia nigra.[11][12]
Myoclonic triangle. The dentate nucleus, along with the red nucleus and the inferior olivary nucleus (ION) of the medulla oblongata, comprises the myoclonic triangle (triangle of Guillain-Mollaret).[1] Via the dentatorubral tract, the dentate nucleus sends fibers through the superior cerebellar peduncle to the contralateral red nucleus. Parvocellular red nucleus axons synapse the ipsilateral ION through the central tegmental tract. Axons of the ION transverse through the inferior cerebral peduncle as climbing fibers to the contralateral cerebellar cortex, synapsing on Purkinje cells. The Purkinje cells synapse the ipsilateral dentate nucleus to complete the myoclonic triangle. Altogether, the myoclonic triangle controls fine voluntary movements.
Cerebellar Purkinje cells. Cerebellar Purkinje cells are GABAnergic neurons located in the cerebellar cortex synapse on deep cerebellar nuclei, such as the dentate nucleus, to inhibit deep cerebellar neurons.[13]
Muscles
The dentate nucleus and the interposed nucleus are involved in modulating voluntary movements of skeletal muscles, especially muscles in the extremities and extraocular muscles.[14]
Physiologic Variants
The cerebellum, and by extension, the dentate nucleus, is highly invariant among humans.[15] Variations usually occur on the arteries supplying the cerebellum.[9]
Surgical Considerations
Resection of tumors in the cerebellum should consider sparing the dentate nucleus as the destruction of the dentate nucleus results in cerebellorubral degeneration with possible hypertrophic ION degeneration and subsequent palatal tremors, which may be caused by the disruption of the myoclonic triangle.[16][1]
Clinical Significance
Cerebellar ataxia. Inactivation of the dentate nucleus results in cerebellar ataxia, a loss of finely coordinated movements.[16] Death of the GABAnergic Purkinje cells, which synapse on the dentate nucleus alongside other deep cerebellar nuclei, can also result in degeneration of the dentate nucleus with subsequent ataxia.[13]
Freiderich ataxia. Friedreich ataxia is the most common inherited autosomal recessive progressive ataxia.[17] Its pathophysiology involves repeating unstable GAA trinucleotides on chromosome 9q with loss of myelination and gliosis in the spinal cord. The cerebellum demonstrates atrophy of the dentate nucleus, possibly due to oxidative stress from a deficiency in frataxin.[18]
Myoclonic triangle disruption. Disruption of the myoclonic triangle, resulting in disinhibition of the ION, can result in palatal tremor, which is associated with rhythmic involuntary contractions of the palatine muscles.[1][19]
Disruption of embryological neural migration. Disruption of neural migration during embryonic development of the cerebellum, such as those that disrupt the reelin pathway or conditions in which reelin expression decreases, can cause a myriad of neuropsychological disorders ranging from congenital ataxia and epilepsy to schizophrenia, mood disorder, and autism.[5]
Maple syrup urine disease. Maple syrup urine disease (MSUD) is a genetic disorder involving the decreased function of the branched-chain alpha-ketoacid dehydrogenase, which is involved in the catabolism of branched-chain amino acids (BCAAs) such as leucine, isoleucine, and valine.[20] Symptoms of MSUD are associated with cerebellar dysfunction, including ataxia and choreoathetosis. Patients with MSUD have a build-up of BCAAs and have a disruption of glutamate synthesis, which is important in learning and memory. Additionally, the build-up of leucine produces cytotoxicity. The magnetic resonance imaging (MRI) and contrast tomography (CT) imaging performed on patients with untreated MSUD will show marked pathognomonic cerebellar and brainstem edema.[21]
Canavan disease. Canavan disease (CD) is a rare early childhood progressive neurodegenerative disease associated with swelling and spongy degeneration of white matter in the brain.[22] There is subsequent atrophy of the cerebellum. Patients with CD have a defective ASPA gene, which produces aspartoacylase. Aspartoacylase is the enzyme that hydrolyzes N-acetylaspartate into aspartate and acetate and is highly expressed in the brain. Deficient aspartoacylase activity allows for the build-up of N-acetyl aspartate, which interferes with myelination of cerebellar neurons in a poorly understood manner.
Langerhans cell histiocytosis. Langerhans cell histiocytosis (LCH) is cancer resulting from the dendritic cells of the skin (Langerhans cell). Although LCH usually affects the skin and bone, it can affect the central nervous system (CNS). T1-weighted MRIs of children with LCH shows changes in the CNS, ranging from pineal gland enlargement to dentate nucleus signaling modifications that may correlate with degeneration.[23]
Alzheimer disease. Normal aging of the cerebellum correlates with a loss of about 2.5% Purkinje cells per decade. However, there is no evidence of age-related neuronal loss of the deep cerebellar nuclei. Patients with Alzheimer's disease have deposits of amyloid plaques on the cerebellar cortex; those at a later stage tend to have balance and gait problems due to a reduction of cerebellar glucose metabolism. Although amyloid and tau inclusions are largely absent from the deep cerebellar nuclei, there is an increased level of iron deposition as per MRI.[24]
Autism spectrum disorders. Autism spectrum disorders are a group of behavioral neurodevelopment disorders associated with cognitive impairment and deficits in language and communication.[25] Changes in cerebellar neuron compositions are related to communicational apraxia observed in children with autism. Several neurophysiological studies examining sectioned brains from children with idiopathic autism have found a reduction in Purkinje cells, especially in the lateral cerebellum. Neurochemical studies determined an alteration in GAD65/67 in Purkinje cells, basket cells, and the dentate nucleus, along with cytoarchitectural changes in the inferior olive and the dentate nucleus; these changes are thought to be associated with asynchrony of Purkinje cell firing. Other findings in autism brains include flocculonodular dysplasia, hypoplastic lobes, and increased neuroglia with glial fibrillary acidic proteins at pathologic sites.
Essential tremor. Even though the thalamic ventral intermediate nucleus has merited consideration as the driving center for tremors, this nucleus receives input from the main cerebellar projections. Research has found that the increased functional connectivity between the cerebellar cortex and dentate nucleus is compensating for the reduced connectivity between the dentate nucleus and thalamus.[26]
Other Issues
Patients undergoing repeated T1-weighted MRI with gadolinium-based contrast agents (GBCAs) for neurologic diseases have a potential for gadolinium to deposit and persist in the globus pallidus and dentate nucleus.[27] This persistence is dose-dependent and depends on cyclization of GBCAs; cyclic GBCAs are more likely to deposit in the globus pallidus and dentate nucleus than linear GBCAs. To date, there is no evidence of adverse health effects from GBCA deposition on the globus pallidus and dentate nucleus. However, manganese-based and iron-based contrast alternatives are undergoing research.
Media
(Click Image to Enlarge)
Myoclonic triangle
Contributed by Katherine Humphreys
(Click Image to Enlarge)
Dentate nucleus
Contributed by Katherine Humphreys
References
Bond KM, Brinjikji W, Eckel LJ, Kallmes DF, McDonald RJ, Carr CM. Dentate Update: Imaging Features of Entities That Affect the Dentate Nucleus. AJNR. American journal of neuroradiology. 2017 Aug:38(8):1467-1474. doi: 10.3174/ajnr.A5138. Epub 2017 Apr 13 [PubMed PMID: 28408628]
Matano S. Brief communication: Proportions of the ventral half of the cerebellar dentate nucleus in humans and great apes. American journal of physical anthropology. 2001 Feb:114(2):163-5 [PubMed PMID: 11169906]
Level 3 (low-level) evidenceBarkovich AJ, Millen KJ, Dobyns WB. A developmental and genetic classification for midbrain-hindbrain malformations. Brain : a journal of neurology. 2009 Dec:132(Pt 12):3199-230. doi: 10.1093/brain/awp247. Epub [PubMed PMID: 19933510]
Butts T, Green MJ, Wingate RJ. Development of the cerebellum: simple steps to make a 'little brain'. Development (Cambridge, England). 2014 Nov:141(21):4031-41. doi: 10.1242/dev.106559. Epub [PubMed PMID: 25336734]
Level 3 (low-level) evidenceRahimi-Balaei M, Bergen H, Kong J, Marzban H. Neuronal Migration During Development of the Cerebellum. Frontiers in cellular neuroscience. 2018:12():484. doi: 10.3389/fncel.2018.00484. Epub 2018 Dec 17 [PubMed PMID: 30618631]
Barthelery NJ, Manfredi JJ. Cerebellum Development and Tumorigenesis: A p53-Centric Perspective. Trends in molecular medicine. 2016 May:22(5):404-413. doi: 10.1016/j.molmed.2016.03.006. Epub 2016 Apr 13 [PubMed PMID: 27085812]
Level 3 (low-level) evidenceLee GH, D'Arcangelo G. New Insights into Reelin-Mediated Signaling Pathways. Frontiers in cellular neuroscience. 2016:10():122. doi: 10.3389/fncel.2016.00122. Epub 2016 May 9 [PubMed PMID: 27242434]
Kim S, Lee H, Lee Y, Lee J, Yang J, Lee M, Yang H. Blood Supply by the Superior Cerebellar Artery and Posterior Inferior Cerebellar Artery to the Motor and Nonmotor Domains of the Human Dentate Nucleus. World neurosurgery. 2019 Feb:122():e606-e611. doi: 10.1016/j.wneu.2018.10.111. Epub 2018 Oct 26 [PubMed PMID: 31108077]
Tubbs RS, Shoja MM, Aggarwal A, Gupta T, Loukas M, Sahni D, Ansari SF, Cohen-Gadol AA. Choroid plexus of the fourth ventricle: Review and anatomic study highlighting anatomical variations. Journal of clinical neuroscience : official journal of the Neurosurgical Society of Australasia. 2016 Apr:26():79-83. doi: 10.1016/j.jocn.2015.10.006. Epub 2015 Dec 7 [PubMed PMID: 26675624]
Sakka L, Coll G, Chazal J. Anatomy and physiology of cerebrospinal fluid. European annals of otorhinolaryngology, head and neck diseases. 2011 Dec:128(6):309-16. doi: 10.1016/j.anorl.2011.03.002. Epub 2011 Nov 18 [PubMed PMID: 22100360]
Dum RP, Strick PL. An unfolded map of the cerebellar dentate nucleus and its projections to the cerebral cortex. Journal of neurophysiology. 2003 Jan:89(1):634-9 [PubMed PMID: 12522208]
Level 3 (low-level) evidenceBostan AC, Strick PL. The basal ganglia and the cerebellum: nodes in an integrated network. Nature reviews. Neuroscience. 2018 Jun:19(6):338-350. doi: 10.1038/s41583-018-0002-7. Epub [PubMed PMID: 29643480]
Koeppen AH. The neuropathology of the adult cerebellum. Handbook of clinical neurology. 2018:154():129-149. doi: 10.1016/B978-0-444-63956-1.00008-4. Epub [PubMed PMID: 29903436]
Soteropoulos DS, Baker SN. Bilateral representation in the deep cerebellar nuclei. The Journal of physiology. 2008 Feb 15:586(4):1117-36. doi: 10.1113/jphysiol.2007.144220. Epub 2008 Jan 10 [PubMed PMID: 18187463]
Level 3 (low-level) evidenceAllen JS, Damasio H, Grabowski TJ. Normal neuroanatomical variation in the human brain: an MRI-volumetric study. American journal of physical anthropology. 2002 Aug:118(4):341-58 [PubMed PMID: 12124914]
Vilis T, Hore J. Effects of changes in mechanical state of limb on cerebellar intention tremor. Journal of neurophysiology. 1977 Sep:40(5):1214-24 [PubMed PMID: 409809]
Level 3 (low-level) evidenceArora R. Imaging spectrum of cerebellar pathologies: a pictorial essay. Polish journal of radiology. 2015:80():142-50. doi: 10.12659/PJR.892878. Epub 2015 Mar 16 [PubMed PMID: 25806100]
Solbach K, Kraff O, Minnerop M, Beck A, Schöls L, Gizewski ER, Ladd ME, Timmann D. Cerebellar pathology in Friedreich's ataxia: atrophied dentate nuclei with normal iron content. NeuroImage. Clinical. 2014:6():93-9. doi: 10.1016/j.nicl.2014.08.018. Epub 2014 Aug 23 [PubMed PMID: 25379420]
Deuschl G, Toro C, Hallett M. Symptomatic and essential palatal tremor. 2. Differences of palatal movements. Movement disorders : official journal of the Movement Disorder Society. 1994 Nov:9(6):676-8 [PubMed PMID: 7845410]
Blackburn PR, Gass JM, Vairo FPE, Farnham KM, Atwal HK, Macklin S, Klee EW, Atwal PS. Maple syrup urine disease: mechanisms and management. The application of clinical genetics. 2017:10():57-66. doi: 10.2147/TACG.S125962. Epub 2017 Sep 6 [PubMed PMID: 28919799]
Steinlin M, Blaser S, Boltshauser E. Cerebellar involvement in metabolic disorders: a pattern-recognition approach. Neuroradiology. 1998 Jun:40(6):347-54 [PubMed PMID: 9689620]
Hoshino H, Kubota M. Canavan disease: clinical features and recent advances in research. Pediatrics international : official journal of the Japan Pediatric Society. 2014 Aug:56(4):477-83. doi: 10.1111/ped.12422. Epub [PubMed PMID: 24977939]
Level 3 (low-level) evidencePorto L, Schöning S, Hattingen E, Sörensen J, Jurcoane A, Lehrnbecher T. Central nervous system imaging in childhood Langerhans cell histiocytosis - a reference center analysis. Radiology and oncology. 2015 Sep:49(3):242-9. doi: 10.1515/raon-2015-0024. Epub 2015 Aug 21 [PubMed PMID: 26401129]
Chen J, Cohen ML, Lerner AJ, Yang Y, Herrup K. DNA damage and cell cycle events implicate cerebellar dentate nucleus neurons as targets of Alzheimer's disease. Molecular neurodegeneration. 2010 Dec 20:5():60. doi: 10.1186/1750-1326-5-60. Epub 2010 Dec 20 [PubMed PMID: 21172027]
Hampson DR, Blatt GJ. Autism spectrum disorders and neuropathology of the cerebellum. Frontiers in neuroscience. 2015:9():420. doi: 10.3389/fnins.2015.00420. Epub 2015 Nov 6 [PubMed PMID: 26594141]
Tikoo S, Pietracupa S, Tommasin S, Bologna M, Petsas N, Bharti K, Berardelli A, Pantano P. Functional disconnection of the dentate nucleus in essential tremor. Journal of neurology. 2020 May:267(5):1358-1367. doi: 10.1007/s00415-020-09711-9. Epub 2020 Jan 23 [PubMed PMID: 31974808]
Smith TE, Steven A, Bagert BA. Gadolinium Deposition in Neurology Clinical Practice. The Ochsner journal. 2019 Spring:19(1):17-25. doi: 10.31486/toj.18.0111. Epub [PubMed PMID: 30983897]